

 inglés
inglés francés
francés español
español ruso
ruso coreano
coreano Japonés japonés
Japonés japonés¿Qué es el polvo de ácido hialurónico de alto peso Molecular?
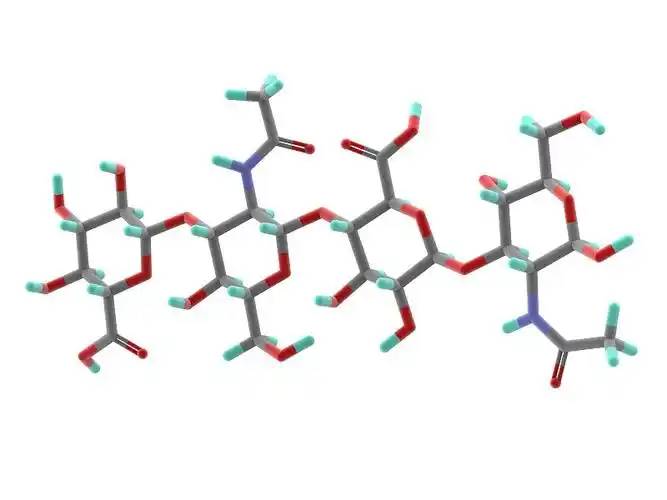
1ácido hialurónico y composición estructural
Hyaluronic Ácido ácido ácido ácido ácido ácido ácido ácido ácido ácido ácido ácido ácido ácido(HA) is a non-sulfated, high molecular weight linear polysaccharide with a basic structure consisting dea disaccharide unit of D-glucuronic acid yN-acetylglucosamine, linked by a β- 1,3- ligand bond between elD-glucuronic acid and the N-acetylglucosamine, and by a β- 1,4- ligand bond between the disaccharide units (Figure 1). The molecular weight of natural hialurónicaacid contained enmost tissues ranges from 1000 to 10 000 kDa, and the definition of hyaluronic acid size has not yetbeen fully agreed upon.According to Naro [1], high molecular weight hyaluronic acid is defined as consisting of more than 500 basic estructuralunits; intermediate molecular weights are generally in the range of 200-500 kDa, and small molecular weight hyaluronic acid is generally defined as less than 200 kDa. According to Monslow etal [2], high molecular weight (HMW hyaluronan) is >1000 kDa, medium molecular weight (MMW hyaluronan) is 250-1000 kDa, low molecular weight (LMW hyaluronan) is >10-250 kDa, and oligo hyaluronan (oligo hyaluronan) is <10 kDa.
The role of hyaluronan in cell motility, cell adhesion and proliferation is mainly mediated by two hyaluronan receptors, CD44 and R-hyaluronan MM (the receptor for hyaluronan-mediated motility, also called CD168). Unlike other mucopolysaccharides, which are synthesised in the Golgi apparatus, hyaluronan is synthesised in the inner membrane of the plasma membrane of the cell by transmembrane synthetases (hyaluronan S1, hyaluronan S2 and hyaluronan S3). Hyaluronan S1 and S2 are responsible for the synthesis of large molecules of hyaluronan and hyaluronan S3 is responsible for the synthesis of small molecules of hyaluronan (<300 kDa), but hyaluronan S3 is more active than hyaluronan S1 and hyaluronan S2 The synthesised large molecules of hyaluronan are cleaved intosmall molecular weights of hyaluronan by ROS (reactive oxygen radicals) and hyaluronan hydrolytic enzyme, and further hydrolysed into low molecular weights of hyaluronan. Further hydrolysis results in oligo- or oligo-hyaluronic acid. Different molecular weights have different physiological functions, e.g. medium molecular weight hyaluronic acid stimulates cell proliferation, while small molecular weight hyaluronic acid promotes cell migration. See Table 1 [3- 19].
2 progreso biológico del ácido hialurónico
2.1 ácido hialurónico y pérdida de peso
Hyaluronic acid has been approved as a new resource food in China (Ministry of Health Announcement No. 12, 2008), and is used as a food additive and food with health benefits in Korea, as a food additive in Japan, and as a supplement in the United States, Canada, Italy, and Belgium.
Breast milk contains the highest level of hyaluronic acid in the first week of life, which decreases to a stable level over the next two months. In general, a 70 kg adult has about 15 g of hyaluronic acid in his or her body, of which nearly one-third is degraded and synthesised on a daily basis. Hyaluronic acid is taken orally to the Piel de pieland does not accumulate excessively in the body; more than 90% of hyaluronic acid is excreted in the breath or urine [19]. Hyaluronic acid is used for weight loss. When hyaluronic acid was administered to C57BL/6 mice at a dosage of 200 mg/kg for 8 weeks, it was found that hyaluronic acid had a good effect on weight loss, which not only reduced body mass, but also reduced adipose tissue and serum LDL-cholesterol, as well as total cholesterol and leptin, and also reduced adipose tissue hyperplasia, and ameliorated fatty liver. It is therefore hypothesised that the main mechanism of hyaluronic acid weight loss may be through the increase of the peroxisome proliferator-activated receptor PPAR-α and the inhibition of PPAR-γ [20].
2.2 ácido hialurónico y envejecimiento
Hyaluronic acid and hyaluronan-binding proteins are involved in cellular senescence. Researchers have found that hyaluronic acid can heal skin damage caused by UV-B exposure through histological changes and wrinkle indicators in vivo. Moreover, high concentrations of hyaluronic acid significantly affected the expression of collagen, matrix metalloproteinase (MMP-1), interleukin IL-1β and interleukin IL-6 protein factors, but not hyaluronan S-2 and transforming growth factor (TGF-β1). Reduction of proteoglycans leads to an increase in free hyaluronan fragments interacting with CD44 protein and phosphorylation of the extracellular signal-regulated kinase ERK1/2, resulting in premature embryonic fibroblast failure [21].

Inhibition of hyaluronan synthesis by 4-methylumbelliferone or targeting of hyaluronan S-2 by miRNA induces cellular senescence, suggesting that hyaluronan plays a role in skin ageing [22]. Además, hyaluronan concentration is greatly reduced in senescent MSCs and in the cell cycle matrix, mainly due to reduced expression of hyaluronidase [23]. Considering the ability of hyaluronan to inhibit fibroblast senescence induced by oxidative stress in vitro [24]. Thus, hyaluronic acid may be involved in ageing during normal and pathological processes. Further investigation of the role of hyaluronic acid in aging in disease states may shed light on the pathogenesis and aid in the development of therapeutic approaches for these diseases.
2.3 ácido hialurónico y crecimiento y desarrollo
Hyaluronan participates in and promotes cell growth through a variety of signalling pathways. In mice in which the hyaluronan gene hyaluronan s2 was knocked out, the heart and blood vessels developed abnormally, and at 9.5 weeks of development, the lack of an intact endocardial cushion resulted in embryonic lethality (TD Camenisch, 2000). Intraperitoneal injections of hyaluronic acid for 3-8 weeks increased the length of villi and depth of the glandular fossa in the small intestine, the depth of the glandular fossa in the colon, and the proliferation of epithelial cells in both. The opposite results were obtained with PEP-1, a short peptide that prevents the binding of hyaluronan to the receptor. These results suggest that endogenous hyaluronan can regulate normal small bowel and colon growth.
Hyaluronic acid and its binding proteins have been shown to play a role in fibroblast proliferation. Hyaluronic acid promotes TGF-β1-mediated fibroblast proliferation, and LMW hyaluronic acid or hyaluronan oligosacáridosstimulate fibroblast proliferation under a variety of conditions, suggesting that hyaluronic acid plays an important role in tissue fibrosis. Hyaluronic acid fragments induced myofibroblast differentiation, endothelial cell differentiation, and chondrogenic differentiation [25], whereas hyaluronic acid and mouse tumour necrosis factor α-inducible protein 6 inhibited cardiomyocyte differentiation and osteoclast differentiation of humanabone marrow mesenchymal stem cells, respectively. These data suggest that hyaluronan and hyaluronan-binding proteins regulate cell differentiation in a complex manner and that this differentiation may be related to cell type and microenvironment.
2.4 ácido hialurónico e inmunoinflamación
Hyaluronic acid and hyaluronan-binding proteins regulate inflammation and the repair of tissue damage by modulating the infiltration of inflammatory cells, the release of inflammatory cytokines and cell migration. Therefore, hyaluronic acid may act as an immunomodulator in human diseases [26]. Synthetic hyaluronic acid has been found to promote the expression of antimicrobial peptides in intestinal epithelial cells. Oral administration of hyaluronic acid from breast milk induced an increase in mouse defensin HβD2 homologue and MuβD3 via Toll-like receptor 4 (TLR4) and CD44 protein, and the addition of hyaluronic acid cytokines to cultured cells showed that colonic mucosal epithelial cells were more resistant to the enteropathogenic bacterium Salmonella typhimurium. Hyaluronic acid regulates cell proliferation and inflammation.
Las células inflamatorias activlos queratinocipara producir interleucina IL- 6, IL-1 y el factor de necrosis tumoral (TNF- -) a través de la liberación de citocinas y la producción de fragmentos de hialuronlmw, que a su vez aumenta la síntesis de hialuronen en parte a través de la acción de HB-EGF, y la retroalimentación hialuronan contribuye a las respuestas inflamatorias incluyendo la migración de fibroblay la proliferación. El ácido hialurónico ha sido reportado por RMA etAl.S. hyaluronan RMA etal [25] informaron que la curcumina acoplal ácido hialurónico mejoró la proliferación de queratinocitos, redujo el daño oxidativo causado por el peróxido de hidrógeno, y mejoró la migración celular en el sitio de los arañazos.
Hyaluronic acid can be utilised by probiotics. Probiotics can use N-acetyl-D glucosamine, a precursor of hyaluronic acid, as a nutrient, and Kim et al [27] found that Lactobacillus plantarum K8 lysate significantly increased hyaluronic acid secretion from cells. Lactobacillus lysates inhibited the Th2 response induced by interleukin 4 (IL-4)-driven stimulation and increased the Th1 response, and lactobacilli controlling the Th1/Th2 balance contributed to hyaluronic acid induction and the reduction of atopic dermatitis lesions.
Hyaluronic acid and CD44 interact with each other and have the ability to modulate T-cell activation, Th1 production, B-cell activation and regulatory T-cell functions. Recent studies have shown that CD44 functions in the adaptation of regulatory T cells by interacting with galactose agglutinin 9 [28].
Hyaluronic acid and CD44 regulate neutrophil adhesion and recruitment. It is endothelial CD44, not neutrophil CD44, that mediates neutrophil migration, and hyaluronic acid is also capable of promoting the production of inflammatory vesicles in response to injury. In CD44-/- deficient mice, lymphocytes preferentially stayed in lymph nodes and delayed their entry into synovial joints with an inflammatory response compared to wild-type cells. CD44-/- deficient mice die from non-infectious pneumonia injury, a continuous inflammatory response characterised by impaired clearance of apoptotic neutrophils [26]. In addition, hyaluronan fragments can affect dendritic cell maturation, e.g. LWM hyaluronan fragments not only induce dendritic cell maturation but also switch on alloimmunity. In addition, hyaluronan oligosacáridosare potential activators of dendritic cells, allowing small molecule hyaluronan fragments to promote cell migration and subsequent allergic modifications.
Además, TLR-4, el receptor natural para los lipopolisacáridos, es uno de los receptores hialurónicos. TLR-4 activa la proteína del factor de transcripción nuclear NF- kB a través de dos vías: una induce citocinas pro-inflamatorias a través del factor de diferenciación mieloide dependiente MyD88, y la otra es una vía MyD88 no dependiente de MyD88 que aumenta los genes pro-inflamindupor interferón a través de interfertipo I [26].
2.5 ácido hialurónico y cáncer
Hyaluronic acid not only acts as a cellular support and hydrophilic matrix, but also regulates cell adhesion, migration, growth and differentiation (TC Laurent, 1992). These properties have led to the involvement of hyaluronic acid in a number of pathological processes. For example, in cancer, hyaluronic acid forms a protective film on the surface of tumour cells, making them less vulnerable to attack by immune cells. At the same time, tumour cells produce more hyaluronic acid or induce the production of hyaluronic acid by releasing growth factors and cytokines. Moreover, ROS-induced hyaluronan fragments contribute to the efficient expression of hyaluronan (R Stern, 2006). Tumour cells and stromal cells are able to express hyaluronan homologues and produce hyaluronan-containing extracellular matrix, which accumulates in the tumour and pericancerous stromal tissues, thus accelerating metastasis of cancer cells [29].
In addition, hyaluronic acid also regulates fibroblast and tumour invasion (Q Yu, 2000). Transforming growth factor and tyrosine kinase, when activated, regulate hyaluronan-mediated invasive cellular motility [27]. Heterogeneity analysis of probe cells using hyaluronan resulted in the identification of subtypes of infiltrating but slow-growing breast tumours. By analysing fibroblastosisolated from transgenic mice overexpressing hyaluronan S2, we found that the cells had a greater ability to invade the stroma. If the hyaluronan S2 gene is knocked out of mesenchymal cells, the invasive fibroblast phenotype is terminated, the accumulation of myofibroblasts is discharged and the development of lung fibrosis is inhibited [30].
2.6 correlación del ácido hialurónico con células madre
The interaction of hyaluronic acid with stem cells has been studied in haematopoietic stem cells, MSCs and pluripotent adult progenitors, suggesting that hyaluronic acid and its binding proteins may interact with various stem cells through their ability to regulate tissue damage and repair processes. During stem cell differentiation, hyaluronan synthesis was enhanced 13-fold and 24-fold, most likely due to an increase in hyaluronan S2 expression. Hyaluronan is required for the production of haematopoietic cells during human embryonic stem cell differentiation. Knockdown of hyaluronan S2 resulted in inhibition of human embryonic stem cell differentiation. Hyaluronan oligosaccharides increase the stem cell properties of epicutápielcells by regulating integrins. Hyaluronic acid promotes CD44-dependent migration of MSCs CD44 has long been used as a marker for stem cells, including embryonic, mesenchymal, haematopoietic and cancer stem cells. In addition, CD44 may play a role in regulating stem cell function. A recent study has shown that although CD44 is not a master stem cell gene, CD44 contributes to stem cell generation and the homeostasis of stem cells in their ecosystem. Hyaluronic acid is required to regulate the haematopoietic support function of bone marrow helper cells and is involved in haematopoietic site assembly.
Hialuronan and Apoptosis (en inglés)
In bleomycin-induced lung injury, hyaluronic acid protects mouse lung epithelial cells from apoptosis. High molecular weight hyaluronic acid reduces UV-induced apoptosis and inflammation in human epithelial corneal cells. Dermal fibroblasts are resistant to stress-induced apoptosis in the presence of high levels of hyaluronan s2 gene expression in the presence of hyaluronan s1/3 knockout, suggesting that hyaluronan S2 protects dermal fibroblasts from environmental stress-induced apoptosis. For inflammatory cells, hyaluronic acid appears to induce apoptosis. Hyaluronic acid appears to induce apoptosis. Hyaluronic acid induces apoptosis in activated T cells via CD44. LMW hyaluronic acid-TLR4 interaction induces apoptosis in inflammatory neutrophils. In rats, administration of hyaluronic acid with a molecular weight of 1600 kDa significantly reduced smoke-induced neutrophil infiltration, pulmonary oedema, respiratory apoptosis and mucus plugging, suggesting that high-molecular-weight hyaluronic acid may have a potential therapeutic effect on smoking-induced lung injury. In addition, cross-linking of CD44 molecule-specific epitopes rapidly induced neutrophil apoptosis in vitro and inhibited neutrophil-dependent renal injury in rats in vivo.
2.8 papel en la migración e invasión celular
Hyaluronic acid mediates inflammatory cell migration to regulate the inflammatory response and tissue damage.CD44 deficiency leads to increased neutrophil migration and lung injury in E. coli pneumonia in mice. TSG6 is a potent inhibitor of neutrophil migration in an in vivo model of acute inflammation. Exogenous Ácido hialurónico de bajo peso molecular, or LWM hyaluronic acid produced by overexpression of HYAL1, promotes the migration of dendritic cells from the skin and subsequently alters the allergic response in a TLR4-dependent manner. Hyaluronan and hyaluronan-binding proteins play a role in fibroblast migration and smooth muscle cell migration. Abnormal accumulation of hyaluronan matrix promotes fibroblast migration.

Specifically sized hyaluronan oligosaccharides stimulate fibroblast migration and excisional wound repair. both CD44 and HMMR have been shown to have a role in fibroblast migration during tissue injury. In addition, myofibroblast migration can be regulated by hyaluronic acid. These studies suggest a role for hyaluronic acid in tissue injury and fibrosis. Small molecular weight hyaluronic acid35 and high molecular weight hyaluronic acid117 showed significant differences in the migration and invasion of breast cancer 4T-1 and SKBR3 cells. In healthy people, the level of hyaluronic acid in the body is 10-100 μg/L,but in breast cancer patients, the level is as high as 200-300 μg/L, and in advanced breast cancer, it reaches 789-2343 μg/L (EH Cooper, 1988). Compared with small molecular weight hyaluronic acid, high molecular weight hyaluronic acid is able to exert a greater squeezing force on the tumour spheres and prevent the growth of cancer cells. Small molecular weight hyaluronic acid can accelerate the process of metastatic fibrosarcoma cell invasion [9].
3 perspectivas
Over the past decades, hyaluronic acid and its hyaluronan receptor have been intensively investigated, leading to an increasing use of hyaluronic acid. Nevertheless, there are still some problems that need to be solved. The first problem to be solved is the definition of high molecular weight and low molecular weight hyaluronic acid. There is no international consensus on the distinction between high and low molecular weight hyaluronic acid, which poses a problem in elucidating the physiological effects of hyaluronic acid on cells. Secondly, high molecular weight hyaluronic acid controls the normal homeostasis of the body and shows anti-inflammatory and anti-cancer effects, while low molecular weight hyaluronic acid and oligo-hyaluronic acid show pro-inflammatory and pro-cancer effects. If the same receptor is used, but the results are significantly different, the mechanism is still not clear. In addition, how hyaluronic acid precisely regulates the occurrence and development of inflammation and tumour cells; whether oral administration of hyaluronic acid affects the intestinal flora, which in turn affects human health; and how modified hyaluronic acid (e.g., sulphated hyaluronic acid) has a different mechanism of action compared with non-modified hyaluronic acid, need to be further elucidated. The solution of these problems will certainly help to prevent diseases and maximise the use of hyaluronic acid for the treatment of human diseases.
referencia
[ 1]Naor D. Editorial: interacción entre hyaluronic acid and Sus receptores (CD44, RHAMM) regula la actividad de la inflamación y el cáncer [J]. Front Immunol, 2016,7:39.
[2]Monslow J,Govindaraju P, E pura. Hyaluronan- a Funcional y structural Dulce dulce Punto en el microambiente del tejido [J ]. Front Immunol, 2015,6 :231.
[3]Karbownik MS,Nowak JZ. Hyaluronan: towards novel anti- cancer Therapeutics [J] (en inglés). Pharmacol Rep, 2013,65(5):1056 — 1074.
[4]Papakonstantinou E, Roth M,Karakiulakis G. hialuronic acid: a key Molecule in skin aging[J]. Dermatoendocrinol, 2012,4(3): 253- 258.
[5]Kavasi RM,Berdiaki A, Ridaki I, Et Al.Metabolismo del HA en homeostasis cutánea y enfermedad inflam[J]. Food Chem Toxicol, 2017,101:128 — 138.
[6]Zhu Y, Hu J, Yu T, et Al.El ácido hialurónico de alto peso molecular inhila la fibrosis del endometri[J]. Med Sci Monit, 2016,22: 3438- 3445.
[7]Zhao YF, Qiao SP, Shi SL,et al. Modular el microambiente tridimensional con hialuronde de diferentes pesos moleculares altera el comportamiento de invasión celular del cáncer de mama [J]. ACS Appl Mater Interfaces. 2017,9(11):9327- 9338.
[8]Noble NoblePW,Lake FR, Henson PM, et al. La activación del hialuronato de CD44 induce la expresión de un factor de crecimiento similar a la insulina - 1 dependiente del factor de necrosis tumoral - alfa - Mecanismo en Murinemacrophages [J]. J Clin Invest, 1993,91(6):2368- 2377.
[9]Hodge- Dufour J, Noble PW, Horton. El Señor, et Al. inducción De IL- 12 y quimiocpor hialuronrequiere adhesión - dependiente priming de residentes pero no provoca macrófagos [J]. J Immunol, 1997, 159 (5): 2492- 2500.
[10]Ohkawara Y, Tamura G, Iwasaki T, et al. Activación y transformación Factor de crecimiento ing - producción de beta en eosinófilos por hialuronan [J]. Am J Respir Cell Mol Biol, 2000,23(4):444- 451.
[11]Fitzgerald KA, Bowie AG, Skeffington BS, et al. Ras, proteína quinasa C Zeta e I Kappa B quinasas 1 y 2 son efecefecposteriores de CD44 durante la activación de fragmentos de ácido hialurónico NF- Kappa Bby en células de carcinoma T- 24 [J]. JImmunol, 2000,164(4):2053 — 2063.
[12]Slevin M, Kumar (Swami) S, «Gaffney» J. Angiogenic oligosaccharides De hialuronan inducmúltiples vías de señalización que afectan vascular Respuestas mitogéde células endoteliy cicatride heridas [J]. J Biol Chem, 2002,277(43): 41046- 41059.
[13]West DC, Kumar S. Hyaluronan and angigenesis [J]. Ciba Found Symp, 1989,143: 187- 201; Discusión 201- 187,281- 185.
[14]West DC, Kumar S. The effect of hyaluronate and ITS oligosaccha- La proliferación de células endoteliy la integridad de la monocapa [J]. Exp Cell Res, 1989,183(1):179- 196.
[15]Vistejnova L, Safrankova B, Nesporova K, et al. Hialuronan mediado por CD44 de bajo peso molecular inducdependiente de IL- 6 y quimiocinas in human dermal fibroblasts potencipotenci Respuesta inmune innata [J]. Cytokine, 2014, 70(2):97- 103.
[16]Termeer CC, Hennies J, Voith U, et al. oligosacáridos Los hialuronson son potentes activadores de las células dendríticas [J]. Inmunol, 2000,165(4):1863 — 1870.
[17]Pandey MS, Baggenstoss BA, Washburn J, et al. The hyaluronan receptor for endocitosis (en inglés) (HARE) activa la expresión génica mediada por NF- kappaB- en respuesta a 40- 400- kDa, pero no más pequeño o más grande, hialuronans [J]. J Biol Chem, 2013, 288(20):14068 — 14079.
[18]Ghatak S, Misra S, Toole BP. Los oligosacáride hialurónicos inhiel crecimiento independiente del anclaje de las células tumorales al suprimir la vía de supervivencia de las células fosfoinositi3 - quinasa /Akt [J]. J Biol Chem, 2002, 277(41): 38013- 38020.
[19]Oe M, Mitsugi K, Odanaka W, et al. El ácido hialurónico alimentario migr. into the skin of Ratas [J]. Científicos científicos mundo Journal, 2014,2014:378024.
[20]Park BG, Park YS, Park JW, et al. Potencial Anti- obesidad de los fragmentos enzimáticos de hialuronan en la obesidad inducida por una dieta rica en grasas en ratones C57BL/6 [J]. Biochem Biophys Res Commun, 2016,473(1): 290 — 295.
[21]Suwan K, Choocheep K, Hatano S, et al. Versican/PG- M ensambhialuronan en la matriz extracelular e inhicd44 - mediada señalización hacia la senescprematura en fibroblaembrion[J]. J Biol Chem, 2009,284(13): 8596- 8604.
[22]Lompardia SL, Papademetrio DL, Mascaro M, et al. Las líneas celulares leucémicas humanas sintetizan hialuronano para evitar la senescencia y resistir la quimioterapia [J]. Glycobiology, 2013,23(12):1463- 1476.
[23]Jung EM, Kwon O, Kwon KS, et al. Evidencias de correlación be- Entre la expresión reducida de VCAM- 1 y la síntesis de hialuronano durante la senesccelular de células madre mesenquimhumanas [J]. Biochem Biophys Res Commun, 2011,404(1): 463- 469.
[24]Cirillo N, Vicidomini A,McCullough M, et Al. A base de ácido hialurónico El compuesto inhiel fibrobla. Senescencia inducida por estrés oxidativo in vitro y previene la mucositis oral in vivo[J]. J Cell Physiol, 2015,230(7): 1421- 1429.
[25]Sharma M, Sahu K, Singh SP, et al. Actividad curativa de heridas Cumina conjugada a ácido hialurónico: evaluación in vitro e in vivo [J]. Artif Cells Nanomed Biotechnol, 2017,28:1- 9.
[26]Jiang D, Liang J, Noble PW. Hialuronano como regulador inmune en enfermedades humanas [J]. Physiol Rev, 2011,91(1):221- 264.
[27]Kim H, Kim HR, Jeong BJ, Et al. Efectos de la ingestión oral de Lactobacillus plantarum K8 lisados derivados de Kim - chi- sobre la hidratación de la piel. J Microb hyaluronic acidiol Biotechno[J], 2015,25(1):74- 80.
[28]Wu C, Thalhamer T, Franca RF, et al. La interacción de galectina - 9- CD44 mejora la estabilidad y la función de las células T reguladoras adaptativas [J]. Inmunidad, 2014, 41(2):270- 282.
[29]Heldin P, Basu K, Olofsson B, et al. La desregulación de la síntesis, degradación y Unión del hialuronano promueve el cáncer de mama [J]. J Biochem, 2013,154(5): 395 — 408.
[30]Li Y, Jiang D, Liang J, et al. La fibrosis pulmonar grave requiere un examen Fenofibroblasivo sivo regulado por hialuronan y CD44[J]. J Exp Med, 2011, 208(7):1459- 1471.
-
anterior
Estudio sobre el uso del ácido hialurónico para vendajes de heridas
-
siguiente
¿Cuál es el uso del hidrogel de ácido hialurónico en la reparación de tejidos?




